Central Fatigue, and the Role of Neurotransmitters on Reduced Work Output

Summary
Ask any coach about fatigue and it’s likely the Central Nervous System will be mentioned during conversation. With all of the conjecture about how the body copes with fatigue and manages the response to stress, the importance of knowing how neurotransmitters interact in the body. If you are curious to know what is the biology…
Introduction, Central and Chronic Fatigue
Sentient organisms are capable of perceiving the status of their moment-to-moment existence. Humans have the ability to compare and contrast the present moment with past experiences, and to rate the quality of such experiences. Many people exist in a state referred to as “chronic fatigue syndrome,” where they are inescapably held in a condition of feeling reduced energy to perform tasks and receive no joy during consciousness.
Anyone who has exercised to the point where they are reaching towards the upper limits of physiology has perceived acute fatigue, which decreases the ability to maintain a specific intensity level of output. According to Taylor et al. (2016), fatigue is a biological warning that signals an organism to reduce physiological output and move towards a resting state. Fatigue can result in performing a task slower or with less coordination, or in the complete inability to perform a task. Fatigue is objectively known to be involved with the performance of a task when there is an increase in EMG readings associated with that task.
Subjectively, fatigue can be said to be present when the participant perceives muscle pain, overall discomfort, or increased effort to perform a task (Taylor et al., 2016). Taylor et al. (2006) stated that factors leading to reduced performance resulting from fatigue are present at every level of the brain-muscle pathway. Meeusen and De Meirleir (1995) reported that exercise clearly alters neurotransmission, and does so by changing the concentration of different neurotransmitters. Such a change in neurotransmission has a direct impact on fatigue.
Chaudhuri and Behan (2004) stated that mechanical work output is a dependent variable affected by many factors. Internal (limbic system) and external (incentives) sources of motivation; feedback from motor, sensory, and cognitive systems; and environmental factors (external…temperature, internal…homeostatic state), are the primary factors that influence work output. The ability to execute and maintain voluntary activity depends on the smooth flow of afferent, interneuron processing, and efferent nervous activity in the primary sensory and motor systems. Any subsystem involved with the relay of information can contribute to fatigue.
Individuals with normal levels of internal and external motivation, and proper sensory and motor functioning, may still have reductions in work output due to limitations such as endocrine abnormality or autonomic dysfunction. Those who display abnormal levels of exertional fatigue, muscular fatigue, and exercise intolerance are categorized as having chronic fatigue syndrome, and are most likely displaying symptoms of neurological disease that is the result of mutation within the mitochondrial DNA. Central fatigue and chronic fatigue syndromes share many common threads, and are often grouped together within the literature (Chaudhuri and Behan (2004). Chaudhuri and Behan (1998) reported that approximately 80% of patients with chronic fatigue had a previous infection that likely led to presentation. These individuals typically have very low levels of motivation, experience anhedonia, struggle with sleep (apnea and hypoxia may be factors), and suffer from depressive symptoms.
Central and chronic fatigue syndromes share many common threads, and are often grouped together. Share on XAccording to Chaudhuri and Behan (2000), central fatigue syndrome presentation is the result of hypothalamic, pituitary, and diencephalon abnormalities. Barron, Noakes, Levy, and Smith (1985) reported that development of sudden and profound central fatigue in athletes due to overtraining involves hypothalamic and neuroendocrine factors. In hypothalamic-pituitary driven cases of central fatigue, changes in body weight and sleep pattern are typically present. Often, centralized fatigue syndromes are the result of diseases that affect the basal ganglia and connected circuitry of the amygdala, thalamus, and frontal cortex. Typically, the connection between the prefrontal cortex and the thalamus is disturbed in these conditions.
Bruno, Crenidge, and Fick (1998) reported that in post-viral central fatigue circumstances, damage will take place in dopaminergic pathways, the reticular activating system, the midbrain, the brainstem, lenticular nuclei, the basal ganglia, thalamus, hypothalamus, and cortical motor areas. Reductions in leptins, substance P, and prostaglandins are associated with hypothalamic-pituitary central fatigue. Fatigue levels can also be impacted by circulating proinflammatory cytokines. These cytokines can be activated when there is a decrease in corticotropin-releasing factor and decreased circulating cortisol.
According to Steinman (1993), acute stress seems to be helpful, based on the fact that there will a spike in cortcotropin-releasing factor, which has an antagonistic effect on the T-helper-1 cell response (a proinflammatory cytokine); however, prolonged chronic stress seems to downregulate this system. This system is downregulated in patients with chronic fatigue syndrome (Scott and Dinan, 1999), post-traumatic stress disorder (Yehuda, 2002), fibromyalgia (McEwan, 1998), and postpolyomyelitis (Bruno, Sapolsky, Zimmerman, 1995). According to Yehuda et al. (2002), low cortisol concentrations following significant psychological or physical trauma were predictive of post-traumatic stress disorder. Low cortisol levels ultimately could impact the glucocorticoid receptors within the hypothalamic-pituitary network by increasing binding sensitivity. Such a change in receptor sensitivity could heighten a state of constant vigilance and feed into a constant stress response.
According to Buckwald, Herreld, and Ashton (2001), the concordance rate among monozygotic twins for the development of centralized fatigue syndrome symptoms was approximately 50%. This suggests that a genetic element, along with environmental factors, is at play in development of this condition. Therefore, certain individuals would be more at risk in terms of moving into a chronically fatigued state due to stress and other environmental influences (e.g., infection).
In susceptible individuals, environmental stressors will cause changes in the hypothalamic-pituitary-adrenal axis and the norepinephrine system. Shannon, Flattem, Jordan et al. (2000) reported that depleted norepinephrine levels, or decreased sensitivity of the receptor for norepinephrine, results in fatigue and depression in animals. Chaudhuri and Behan (2004) stated that organisms have opposing directional responses regarding development of fatigue with acute versus chronic stress experience, where chronic stress moves animals towards chronic fatigue syndromes.
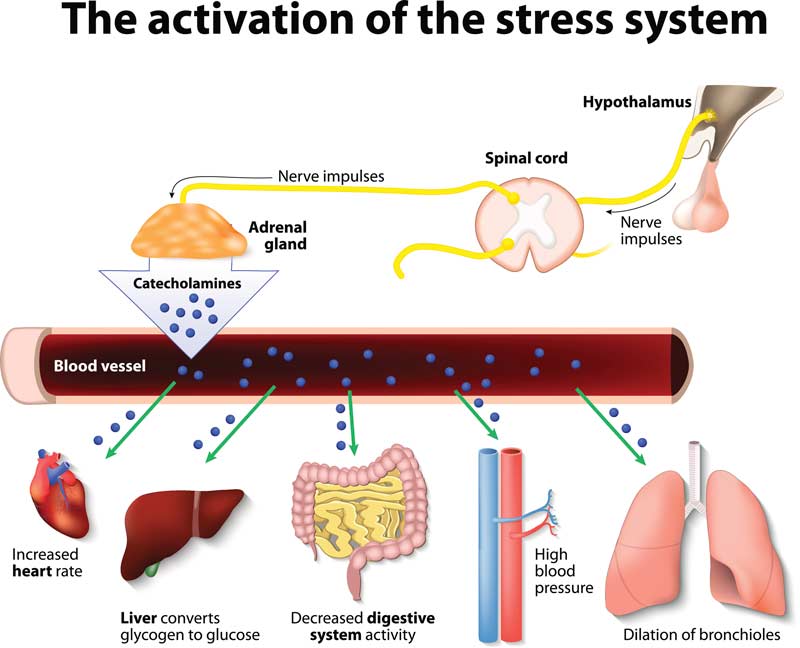
Changes in synaptic receptor sensitivities to corticotropin-releasing factor, serotonin, and norepinephrine establish the nature and severity of the fatigue experience. In humans, prolactin is also secreted under stressful circumstances from the anterior hypothalamus. Dopamine is known to have an antagonistic effect on prolactin secretion. This has led some to believe that dopamine may have a role in combatting elements of the chronic fatigue response system (Chaudhuri and Behan, 2004).
Central Fatigue from an Acute Work Perspective, Examining Neurotransmitters
Newsholme et al. (1987) were the first to report on the notion of central fatigue within acute mechanical work circumstances. They stated that central fatigue was a serotonin-mediated phenomenon, where rising concentrations of serotonin led to an increased perception of lethargy, sleepiness, and reduced motivation. The subsequent investigation has led to mixed results in validating the serotonin-centric hypothesis of central fatigue. However, the consensus scientific perspective points to neurotransmitter concentrations and specific receptor binding activity in specific parts of the brain as being the driver of demonstrable organism fatigue resulting from central pathways. The following section will analyze the findings within the literature on the impact of serotonin, norepinephrine, and dopamine on central fatigue in acute work output.
Neurotransmitters are the chemical messengers that relay information from one neuron to the next within the central nervous system. In regards to acute central fatigue models, monoamine neurotransmitters, which include serotonin, norepinephrine, and dopamine, are believed to be the key players. According to the meta-analysis done by Taylor et al. (2016), the result of central fatigue is either decreased voluntary muscle force or maintenance of the same muscle force via a compensatory strategy somewhere in the neural system.
The primary factor leading to central fatigue is an exercise-induced alteration in monoamine neurotransmitter concentration in specific parts of the brain. The most difficult factor in determining the exact role of the monoamine neurotransmitters on fatigue is that each monoamine causes different responses depending on which region of the brain the binding to receptors is taking place in. To provide clarity on this topic, it becomes prudent to analyze each neurotransmitter and determine what the critical regions of the brain are for receptor binding on fatigue.
Serotonin
Perhaps the two most threatening environmental factors on survival of an organism are temperature and pH. Humans belong to the category of animals known as homeotherms, which have to maintain a relatively constant body temperature, often via internal heat production. Regarding monoamine neurotransmitters and internal heat production, Soares, Coimbra, and Marubayashi (2007) found that increased concentration of serotonin in the preoptic area is associated with greater heat production during exercise. Newsholme et al. (1987) first reported that central fatigue was a serotonin-based phenomenon. Newsholme, Blomstrand, and Ekblom (1992) claimed that the mechanism of central fatigue was elevated tryptophan levels that led to increased serotonin concentrations, and that serotonin precipitated feelings of lethargy and reduction of motivation.
Soares, Lima and Coimbra (2003), and Soares, Lima, and Coimbra (2004) found that increased hypothalamic tryptophan levels precipitated fatigue and were related to a rise in core temperature marked by increased internal heat production and decreased ability to dissipate heat to the environment. Gisolfi and Moura (2000) and Zhang et al. (1997) found that dissipation of heat from the body is more important for temperature regulation compared to heat production during exercise conditions. Nielsen et al. (1997), and Walters et al. (2000) reported that CNS drive towards exercise work output was reduced by elevated core temperature, and that hyperthermia led to subjective discomfort and lethargy. This decrease in CNS outflow and motivation was believed to be a safeguard against allowing dangerously high brain temperatures to occur. Rodrigues et al. (2003) showed that heat storage rates in rats was the main limiting factor in running performance with thermo-neutral and hot environments.
Coimbra and Migliarini (1986), Ferreira, Marubayashi, and Coimbra (1999), Lin et al. (1998), and Santos, Leite, and Coimbra (1991) all determined that the preoptic area and the anterior hypothalamus are the two parts of the brain most responsible for thermoregulation. These parts of the brain are also critical for evaluating and regulating external thermal inputs with metabolically produced heat. Lin et al. (1998) found that injecting serotonin into the preoptic area and hypothalamus of rats increased core body temperature. Soares, Coimbra, and Marubayashi (2007) reported that rats injected with tryptophan into the right cerebral ventricle showed decreased running performance along with increased core temperature and increased concentrations of serotonin in the preoptic area and hypothalamus, compared to rats injected with saline (control condition).
Furthermore, Soares, Coimbra, and Marubayashi (2007) showed that serotonin concentrations were also increased in the hippocampus of rats injected with tryptophan, compared to control. Running time to fatigue was directly correlated with serotonin concentrations in the hippocampus, through a mechanism that seemed to have nothing to do with hyperthermia. Overall, a fatiguing mechanistic cascade appears to exist between tryptophan levels in the brain, which act as a precursor to serotonin, which binds to the preoptic area and the hypothalamus for a heat production element inducing fatigue, as well as a separate serotonin effect on the hippocampus, which has some other fatigue-inducing response. Serotonin concentrations in these areas appear to have a linear relationship with the onset of fatigue.
Serotonin concentrations in these areas appear to have a linear relationship with fatigue onset. Share on XRegarding the mechanism by which serotonin concentrations in the hippocampus impact fatigue, conclusive evidence still remains elusive. Meeusen et al. (1996), Takahashi et al. (2000), and Wilson and Marsden (1996) found a relationship between serotonin levels in the hippocampus and locomotion. Serotonergic neurons that descend from the hindbrain to the spinal cord appear to be involved in central pattern generator (CPG) neural activity controlling locomotion. Without serotonin being present in these neurons, locomotion capabilities are lost.
Soares, Coimbra, and Marubayashi (2007) found that rising concentrations of serotonin in hippocampal neurons lead to a linear increase of fatigue in running rats, and reported that mechanisms need further elaboration. Soares, Coimbra, and Marubayashi (2007) reported that their findings call into question which neurons associated with the serotonergic system are responsible for fatigue. Perhaps there is interplay between multiple systems; perhaps only one set is truly responsible. Consideration of the fact that other neurotransmitters in other parts of the brain may be modifying these effects as well must be considered.
Sharples, Koblinger, Humphreys, and Whelan (2014) reported that monoamines promote locomotion and influence the rhythmicity of locomotion. This influence occurs via binding to the corticospinal tract as well as hind brain regions, and motoneurons. Cotel, Exley, Cragg, and Perrier (2013) found that serotonin contributes to fatigue primarily through binding to motoneurons. According to Johnson et al. (2004), and Perrier, Rasmussen, Christensen, and Petersen (2013), synapses from descending tracts of serotonergic neurons are adjacent to the dendrites and cell bodies of motoneurons. Serotonin binds to the 5HT2 and 5HT1A receptors in the motoneuron system. 5HT2 receptors are excitatory and appear to be the receptor that allows serotonin to be involved with promoting locomotion. When serotonin levels reach very high levels, a spillover effect will be seen, and serotonin will begin binding to 5HT1A receptors (which are known to be inhibitory) and will prevent motoneuron firing.
When examining the effects of serotonin on fatigue from the perspective of the motoneuron, it appears as though there are two distinct ways in which this occurs. Fornal, Martin-Cora, and Jacobs (2006) showed that, in cats, there is an eventual decrease in concentrations of serotonin with prolonged exercise (removal of the excitatory stimulus at the dendritic binding site). Wei, Glaser, and Deng (2014), through indirect measures with humans, found that serotonin release increases in concentration as the force of muscular contraction increases in exercise (addition of the inhibitory axonal binding site). No studies on humans have directly measured serotonin concentrations in the brain; however, 5HT1A receptors are known to exist in humans, and D’Amico et al. (2015) showed that motoneuron excitability in humans can be reduced via 5HT1A agonist drugs administration.
Dopamine
The original hypothesis on central fatigue related primarily to the effects exerted by serotonin on the system; however, researchers also began to understand that other monoamines were powerful players in regards to fatigue. In 1972, Borg et. al, showed that administration of amphetamines improved performance during exercise. Bailey et al. (1993) demonstrated the importance of increased concentrations and binding of dopamine during exercise. Fatigue in rats was correlated with increased serotonin and reduced dopamine in the brain stem and midbrain. Davis and Bailey (1997) showed that the interaction between serotonin and dopamine influenced central nervous system fatigue. A low ratio of serotonin to dopamine favors improved performance and a high ratio decreases motivation and promotes lethargy, resulting in decreased performance (Davis & Bailey, 1997).
Researchers began to understand that other monoamines were powerful players in regards to fatigue. Share on XEarly studies where researchers administered amphetamines showed significant increases in performance in both animals (Gerald, 1978; Heyes et al., 1985) and humans (Wyndham et al., 1971; Borg, 1972). Watson et al. (2005) investigated the effects of bupropion, a dual dopamine and norepinephrine reuptake inhibitor, during cycling at 18 and 30.1 degrees Celsius. At 18 degrees Celsius, no difference was found between the placebo and bupropion trial. When subjects cycled at 30.1 degrees Celsius, the bupropion group performed 9% faster. Roelands, Hasegawa, Watson et al. (2008) applied the same cycling and temperature protocol and administered methylphenidate to our subjects. At 18 degrees Celsius, no difference in performance was observed between conditions. Methylphenidate administration improved cycling time trial performance in 30 degrees Celsius by 16% compared to placebo. The subjects receiving methylphenidate were able to reach significantly higher core temperatures compared to controls (40.1 vs 39.1 respectively).
Despite reaching significantly higher core temperatures, the experimental subjects reported the same subjective perception of thermal sensation and rating of perceived exertion (RPE) as control subjects. The researchers concluded that the increase in dopamine concentration has an effect on the internal safety switch of the brain regarding thermoregulation. The increased dopamine led to a state where the subjects ignored the potentially harmful effects of increased temperature as well as enhanced power production.
Foley and Fleshner (2008) stated that the reason that you’ll see improved performance in hot environments versus no change in ambient temperatures with dopamine administration is that enhanced dopamine levels will provide an increase in psychological motivation to work harder in a more stressful environment. Increased dopamine levels will offset the negative effects on motivation that are brought on by the stress of heat exposure (Del Arco & Mora, 2009). The mechanism of the ergogenic effect of increased dopamine concentrations is believed to be the result of stimulation of the ventral tegmental area. According to Burgess et al. (1991), rats that received stimulation to the ventral tegmental area demonstrated increased motivation to run on a treadmill compared to rats receiving an electric shock.
Norepinephrine
Roelands et al. (2013) reported that different neurotransmitter concentrations in the brain can lead to different pacing strategies during cycling time trial performance. Subjects with higher concentrations of dopamine in the brain demonstrated higher power output throughout time trial performance compared to control. Conversely, increased serotonin and norepinephrine led to decreased power output during time trial performance. The researchers stated that with increased serotonin, subjects were unable to end the time trial with an extra surge of power output (a kick). This led the researchers to speculate that serotonin may cut off access to some type of reserve capacity of substrate or perhaps decreases the motivation to increase power output (Roelands et al., 2013).
Serotonin, dopamine, and norepinephrine-dominated neuronal tracts innervate different areas of the hypothalamus, including the preoptic and anterior hypothalamus, which are considered the most critical regions of the brain for thermoregulation monitoring and control responses. Quan et al. (1991, 1992) have shown administering norepinephrine into the preoptic area of conscious guinea pigs decreased core temperature via reduced metabolic rate. Roelands et al. (2008) showed that subjects cycled 20% longer after given reboxitane, a norepinephrine reuptake inhibitor, to complete the same amount of work in comparison with the placebo trial. Subjects reported feeling colder before and during exercise under the reboxitane condition, and core temperature tended to be lower during exercise.
When studying the effects of norepinephrine on fatigue, part of the difficulty in the process relates to the fact that administration of drugs, such as reboxitane, exerts simultaneous central (brain) and peripheral (heart and vasculature) effects. Such combined effects can lead to confounds in elucidating the exact central role that norepinephrine plays (increased heart rate could expedite the experience of fatigue). Klass, Roelends, Levenez et al. (2012), Piacentini, Meeusen, Buyse et al. (2002), and Roelends, Goekint, Heyman et al. (2008) have all reported that norepinephrine reuptake inhibitor drugs have shown no effect or negative effect on reducing fatigue during exercise conditions with humans.
The negative effect on performance associated with pharmacologically increased norepinephrine levels is unexpected because of the positive associations between norepinephrine and levels of arousal and sensations of reward (Montgomery, 1997; Meeusen et al., 2006). Actions of norepinephrine on neurons alters serotonergic activity via binding to excitatory a1-adrenoceptors. Activation of the a1-adrenoceptors in the locus coeruleus causes action potentials in the serotonergic neurons in the brain stem and dorsal raphe (Szabo & Blier, 2001). Based on these interactions, the negative effects of norepinephrine on fatigue may be modulated through the serotonergic system of the brain.
Conclusions
Fatigue is a perceived experience that is unpleasant and is associated with decreased performance in physical tasks. Fatigue is induced by situations that are associated with a heightened stress experience. Elevated stress experiences that are short term stimulate organisms and have the ability to improve underlying physiology, and resiliency to future stressors. Stress experiences that are prolonged have deleterious effects on physiological systems. When animals exist in ceaseless stressful environments, they have the potential to develop chronic fatigue syndrome that has a stereotypical subjective component associated with lethargy and the inability to experience joy, as well as a chemical profile objectively linked to altered glucocorticoid levels and receptor abnormalities.
Exercise presents a specific environmental condition wherein the participant moves into an acutely fatigued state that results in the inability to sustain an absolute level of mechanical work output. Chronic and acute physiological state differences present themselves in the examination of many different systems, and are often extremely divergent from one another in terms of desirability on the impact of the overall status of an organism. Despite such differences, some of the mechanisms that lead to an overall state, such as fatigue, whether it be chronic or acute in nature, may find a common denominator.
From a reductionist perspective, central fatigue, whether chronic or acute, appears to involve specific neuronal circuitry, which is impacted by the concentration of specific neurotransmitters in the synapse as well as the binding sensitivity with receptors. While it is extremely early in the examination of the impact of neurotransmitters on fatigue, there appears to be a relationship with monoamine concentrations and perception of fatigue. Maintaining a situation where dopamine concentrations are protected and do not drop to diminished levels appears to be an effective strategy for preventing the movement towards a fatigued state.
Short-term stimulatory activity has the potential to increase dopamine levels. Dopamine appears to have a neuroprotective role against creating an unfavorable glucocorticoid level and aberrant receptor activity situation. Furthermore, dopamine acts in a rewarding manner, and reinforces the behavior that resulted in the initial secretion, thus making it likely that the behavior will be executed again. While this relationship of fatigue resistance and dopamine concentrations appears to be emerging based on the current body of literature, patience and skepticism should guide the overall thought-process regarding causative roles. Behavioral outcomes from neuronal activity driven through neurotransmitter communication systems are modulated by the interaction of many neurotransmitters working together in a concert-like fashion.
We are in the age of the brain in the body of knowledge of exercise physiology. However, this age is in its infancy, and our understanding is both superficial and lacking an overall integrated framework to guide our perspective on individual study findings. Future findings will be transformative in the way that we view chronic and acute states of fatigue, and new methods for manipulating the system will likely present themselves that lead to breakthroughs in physiological output.
References
- Barron JL, Noakes TD, Levy W, Smith C. Hypothalamic dysfunction in overtrained athletes. J Clin Endocrinol Metab 1985; 60: 803–06.
- Borg G, Edstrom CG, Linderholm H, et al. Changes in physical performance induced by amphetamine and amobarbital. Psychopharmacologia (1972): 26(1): 10–18.
- Bruno RL, Crenage SJ, Fick NM. Parallels between post-polio fatigue and chronic fatigue syndrome: a common pathophysiology? Am J Med (1998); 105: 66S-73S.
- Bruno RL, Sapolsky R, Zimmerman JR, Fick NM. The pathophysiology of a central cause of post-polio fatigue. Ann N Y Acad Sci (1995); 753: 257–75.
- Buchwald D, Herrell R, Ashton S, et al. A twin study of chronic fatigue. Psychosom Med (2001); 63: 936–43.
- Burgess ML, Davis JM, Borg TK, Buggy J. Intracranial self-stimulation motivates treadmill running in rats. J Appl Physiol (1991)S: 71(4): 1593–1597.
- Chaudhuri A, Behan PO. Neurological dysfunction in chronic fatigue syndrome. J Chron Fatigue Syndr (2000); 6: 51–68
- Chaudhuri A, Behan PO, Fatigue in neurological disorders. The Lancet (2004); 363: 978–88
- Chaudhuri A, Behan WMH, Behan PO. Chronic fatigue syndrome. Proc R Coll Physicians Edinb (1998); 28: 150–63.
- Coimbra CC, Migliorini RH, Insulin-sensitive glucoreceptors in rat preoptic area that regulate FFA mobilization, Am. J. Physiol. 251 (1986) E703–E706.
- Cotel F, Exley R, Cragg SJ, Perrier JF. Serotonin spillover onto the axon initial segment of motoneurons induces central fatigue by inhibiting action potential initiation. Proc Natl Acad Sci U S A. (2013);110(12):4774–9.
- D’Amico JM, Butler AA, Butler JE, Gandevia SC, Taylor JL. Activation of 5HT1A receptors: a plausible contributor to central fatigue. Malmo, Sweden: In: Congress of the European College of Sport Science; (2015).
- Davis JM, Bailey SP. Possible mechanisms of central nervous system fatigue during exercise. Med Sci Sports Exerc (1997): 29(1): 45–57.
- Del Arco A, Mora F. Neurotransmitters and prefrontal cortex–limbic system interactions: implications for plasticity and psychiatric disorders. J Neural Transm (2009): 116: 941–952.
- Ferreira ML, Marubayashi U, Coimbra CC, The medial preoptic area modulates the increase in plasma glucose and free fatty acid mobilization induced by acute cold exposure, Brain Res. Bull. 49 (1999) 189–193.
- Foley T, Fleshner M. Neuroplasticity of dopamine circuits after exercise: implications for central fatigue. Neuromol Med (2008): 10: 67–80
- Fornal CA, Martin-Cora FJ, Jacobs BL. ‘‘Fatigue’’ of medullary but not mesencephalic raphe serotonergic neurons during locomotion in cats. Brain Res. (2006);1072(1):55–61.
- Gerald MC. Effects of (1)-amphetamine on the treadmill endurance performance of rats. Neuropharmacology (1978): 17(9): 703–704.
- Gisolfi CV, Moura F, What’s so important about a body temperature of 37 ◦C? in: Gisolfi CC, Moura F (Eds.), From The hot brain: Survival temperature, and human body, MIT Press, Cambridge, MA, (2000), pp. 95–119.
- Gonzalez-Alonso J, Teller C, Andersen SL, Jensen FB, Hyldig T, Nielsen B, Influence of body temperature on the development of fatigue during prolonged exercise in the heat J. Appl. Physiol. 86 (1999) 1032–1039
- Harbuz MS, Rees RG, Ecland D, et al. Paradoxical responses of hypothalamic CRF mRNA and CRF-41 peptide and adenohypophyseal POMC mRNA during chronic inflammatory stress. Endocrinology (1992); 130: 1394–400.
- Heyes MP, Garnett ES, Coates G. Central dopaminergic activity influences rats ability to exercise. Life Sci (1985): 36(7): 671–677.
- Klass M, Roelands B, Le´ve´nez M, et al. Effects of noradrenaline and dopamine on supraspinal fatigue in well-trained men. Med Sci Sports Exerc. (2012);44(12):2299–308.
- Johnson KV, Edwards SC, Van Tongeren C, Bawa P. Properties of human motor units after prolonged activity at a constant firing rate. Exp Brain Res. (2004);154(4):479–87.
- Lin MT, Tsay HJ, Su WH, Chueh FY, Changes in extracellular serotonin in rat hypothalamus affect thermoregulatory function, Am. J. Physiol. 274 (1998) R1260–R1267.
- McEwen BS. Protective and damaging effects of stress mediators. N Engl J Med (1998); 338: 171–79.
- Meeusen R, De Meirleir K. Exercise and Brain neurotransmission. Sports Med (1995): 20(3): 160–188.
- Meeusen R, Roelands B, Central fatigue and neurotransmitters, can thermoregulation be manipulated? Scandinavian Journal of Medicine and Science in Sports (2010) 20(3) 19-28.
- Meeusen R, Thorre K, Chaouloff F, Sarre S, De Meirleir K, Ebinger G, Michotte Y, Effects of tryptophan and/or acute running on extracellular 5-HT and 5-HIAA levels in the hippocampus of food-deprived rats, Brain Res. 740 (1996) 245–252.
- Meeusen R, Watson P, Dvorak J. The brain and fatigue: new opportunities for nutritional interventions? J Sports Sci (2006a): 24: 773–782.
- Meeusen R, Watson P, Hasegawa , et al. Central fatigue. The serotonin hypothesis and beyond. Sports Med (2006b): 36(10): 881–909.
- Montgomery S. Reboxetine: additional benefits to the depressed patient. J Psychopharmacol (1997): 11: S9–S15.
- Newsholme EA, Acworth I, Blomstrand E. Amino acids, brain neurotransmitters and a functional link between muscle and brain that is important in sustained exercise. In: Benzi G, editor. Advances in Myochemistry. London: John Libbey Eurotext; (1987). pp. 127–33.
- Newsholme EQ, Blomstrand E, Ekblom B, Physical and mental fatigue: metabolic mechanisms and importance of plasma amino acids, Br. Med. Bull. 48 (1992) 477–495.
- Nielsen B, Strange S, Christensen NJ, Warberg J, Saltin B, Acute and adaptative responses in human to exercise in warm humid environment, Pflugers Arch. 43 (1997) 49–56.
- Perrier JF, Rasmussen HB, Christensen RK, Petersen AV. Modulation of the intrinsic properties of motoneurons by serotonin. Curr Pharm Des. (2013);19(24):4371–84.
- Piacentini MF, Meeusen R, Buyse L, et al. No effect of a noradrenergic reuptake inhibitor on performance in trained cyclists. Med Sci Sports Exerc. (2002);34(7):1189–93.
- Quan N, Xin L, Blatteis C. Microdialysis of norepinephrine into preoptic area of guinea pigs: characteristics of hypothermic effect. Am J Physiol (1991): 261: R378–R385. Central fatigue 27
- Quan N, Xin L, Ungar A, et al. Preoptic norepinephrine-induced hypothermia is mediated by alpha 2-adrenoceptors. Am J Physiol (1992): 262: R407–R411.
- Rodrigues LOC, Oliveira A, Lima NRV, Machado-Moreira CA, Heat storage rate and acute fatigue in rats, Braz. J. Med. Biol. Res. 36 (2003) 131–135.
- Roelands B, de Koning J, Foster C, Hettinga F, Meeusen R. Neurophysiological determinants of theoretical concepts and mechanisms involved in pacing. Sports Med. (2013);43(5):301–11.
- Roelands B, Goekint M, Buyse L, et al. Time trial performance in normal and high ambient temperature: is there a role for 5-HT? Eur J Appl Physiol. (2009);107(1):119–26.
- Roelands B, Goekint M, Heyman E, et al. Acute norepinephrine reuptake inhibition decreases performance in normal and high ambient temperature. J Appl Physiol (1985). (2008);105(1):206–12.
- Roelands B, Hasegawa H, Watson P, et al. The effects of acute dopamine reuptake inhibition on performance. Med Sci Sports Exerc. (2008);40(5):879–85.
- Roelands B, Meeusen R. Alterations in central fatigue by pharmacological manipulations of neurotransmitters in normal and high ambient temperature. Sports Med. (2010);40(3):229–46.
- Santos GL, Leite JVP, Coimbra CC, Metabolic adjustments during adaptation to high ambient temperature by preoptic-lesioned rats, Braz. J. Med. Biol. Res. 24 (1991) 1169–1172.
- Scott LV, Dinan TG. The neuroendocrinology of chronic fatigue syndrome: focus on the hypothalamic-pituitary-adrenal axis. Funct Neurol (1999); 14: 3–11.
- Shannon JR, Flattem NL, Jordan J, et al. Orthostatic intolerance and tachycardia associated with norepinephrine transporter deficiency. N Engl J Med (2000); 342: 541–49.
- Sharples SA, Koblinger K, Humphreys JM, Whelan PJ. Dopamine: a parallel pathway for the modulation of spinal locomotor networks. Front Neural Circuits. (2014);8:55.
- Soares DD, Coimbra CC, Marubayashi U. Tryptophan-induced central fatigue in exercising rats is related to serotonin content in the preoptic area. Neuroscience Letters 415 (2007) 274–278
- Soares DD, Lima NRV, Coimbra CC, Marubayashi U, Intracerebroventricular tryptophan increases heating and heat storage rate in exercising rats, Pharmacol. Biochem. Behav. 78 (2004) 255–261.
- Soares DD, Lima NRV, Coimbra CC, Marubayashi U, Evidence that tryptophan reduces mechanical efficiency and running performance in rats, Pharmacol. Biochem. Behav. 74 (2003) 357–362.
- Steinman L. Autoimmune disease. Sci Am (1993); 269: 106–14.
- Szabo S, Blier P. Functional and pharmacological characterization of the modulatory role of serotonin on the firing activity of locus coeruleus norepinephrine neurons. Brain Res (2001): 922: 9–20.
- Takahashi H, Takada Y, Nagai N, Urano T, Takada H, Serotonergic neurons projecting to hippocampus activate locomotion, Brain Res. 869 (2000) 194–202.
- Taylor JL, Amann M, Duchateau J, Meeusen R, Rice CL. Neural Contributions to Muscle Fatigue: From the Brain to the Muscle and Back Again. Med and Science in Sports and Exercise. (2016); 48(11): 2294–2306.
- Taylor JL, Todd G, Gandevia SC. Evidence for a supraspinal contribution to human muscle fatigue. Clin Exp Pharmacol Physiol (2006): 33: 400–405.
- Walters TJ, Ryan KL, Tate LM, Mason PA, Exercise in the heat is limited by a critical internal temperature, J. Appl. Physiol. 89 (2000) 799–806
- Watson P, Hasegawa H, Roelands B, Piacentini MF, Looverie R, Meeusen R. Acute dopamine/noradrenaline reuptake inhibition enhances human exercise performance in warm, but not temperate conditions. J Physiol. (2005);565(Pt 3):873–83.
- Wei K, Glaser JI, Deng L, et al. Serotonin affects movement gain control in the spinal cord. J Neurosci. (2014);34(38):12690–700.
- Wilson WM, Marsden CA, In vivo measurement of extracellular serotonin in the ventral hippocampus during treadmill running, Behav. Pharmacol. 7 (1996) 101–104.
- Wyndham CH, Rogers GG, Benade AJ, Strydom NB. Physiological effects of the amphetamines during exercise. S Afr Med J (1971): 45(10): 247–252.
- Yehuda R. Current status of cortisol findings in post-traumatic stress disorder. Psychiatr Clin North Am (2002); 25: 341–68.
- Zhang YH, Hosono T, Yanase-Fujiwara TM, Chen XM, Kanosue K, Effect of midbrain stimulations on thermoregulatory vasomotor responses in rats, J. Physiol. 503 (1997) 177–186.

{kind=link}